|



ЭНЦИКЛОПЕДИЯ РАДИОЭЛЕКТРОНИКИ И ЭЛЕКТРОТЕХНИКИ Мощный импульсный стабилизированный блок питания. Энциклопедия радиоэлектроники и электротехники
Энциклопедия радиоэлектроники и электротехники / Блоки питания В предлагаемой вниманию читателей статье описан мощный импульсный источник для питания различной электронной аппаратуры. Он собран по схеме полумостового инвертора, управляемого ШИ-контроллером TL494. Появление мощных высоковольтных полевых транзисторов явилось предпосылкой для развития сетевых высокочастотных блоков питания с широт-но-импульсным (ШИ) управлением [1,2]. Основные преимущества подобных источников перед традиционными линейными - получение большей мощности на нагрузке при меньших габаритах и, соответственно, большего КПД [3]. Схема предлагаемого импульсного блока питания показана на рис. 1. Основа устройства - преобразователь, собранный по полумостовой схеме. В источнике питания выполнена полная гальваническая развязка между входной высоковольтной и выходной цепями. Узел управления собран на основе ШИ-контроллера TL494.
Основные технические характеристики источника питания
Транзисторный оптрон U2 обеспечивает гальваническую развязку в цепи отрицательной обратной связи по напряжению. Падение напряжения на резисторе R7 приблизительно равно 2,5 В. Сопротивление этого резистора рассчитывают, задавшись током через резистивный делитель R6R7. Сопротивление резистора R6 вычисляют по формуле
где Uвыx - выходное напряжение источника питания; I1 - ток через резистивный делитель R6R7. Сопротивление резистора R9 определяет ток через излучающий диод оптрона U2.1, а также минимальный рабочий ток стабилизатора DA1. При выбранном токе в этой цепи I2 (значение тока должно быть в допустимых пределах для стабилизатора DA1) сопротивление резистора R9 рассчитывают по формуле
где U F - падение напряжения на излучающем диоде оптрона U2.1. Микросхема DA5 стабилизирует напряжение 8 В для питания делителя, состоящего из фототранзистора оптрона U2.2 и резистора R17. Напряжение от средней точки делителя поступает на неинвертирующий вход первого усилителя сигнала ошибки ШИ-контроллера DA6. Напряжение для питания узла управления и драйверов (микросхема DA7) полевых транзисторов обеспечивает вспомогательный источник на сетевом трансформаторе Т2 и аналоговых стабилизаторах напряжения DA2 и DA3. Узел защиты по току собран на компараторе DA4 и триггере DD1.1. Функцию датчика тока выполняет резистор R5, включенный в диагональ полумоста. На неинвертирующий вход компаратора DA4 подается напряжение треугольной формы с конденсатора (С26) частотозадающей цепи тактового генератора ШИ-контроллера (рис. 2). На выходе компаратора формируются тактовые импульсы, поступающие на вход С триггера DD1.1.
Если падение напряжения на резисторе R5 достигнет 1,1В, включаются излучающие диоды и открывается фототранзистор оптрона U1. На вход S триггера DD1.1 поступит низкий уровень. На прямом выходе триггера DD1.1 и, следовательно, на неинвертирующем входе второго усилителя сигнала ошибки ШИ-контроллера DA6 установится высокий уровень. В этом случае оба транзистора VT1 и VT2 будут закрыты. Для управления мощными коммутирующими полевыми транзисторами применена специализированная микросхема - двуканальный драйвер DA7. На рис. 3 показана внутренняя структура одного канала. В скобках указаны номера выводов второго канала. Каждый канал содержит оптрон и усилитель с мощным токовым выходом. Подобные микросхемы широко используют для управления как асинхронными, так и электродвигателями постоянного тока. Параметры драйвера позволяют непосредственно управлять полевыми транзисторами с изолированным затвором, коммутирующими ток до 50 А при напряжении не более 1200 В. Основные параметры микросхемы HCPL315J
Сопротивление резисторов R3 и R4 в цепях затворов коммутирующих транзисторов рассчитывают по формуле
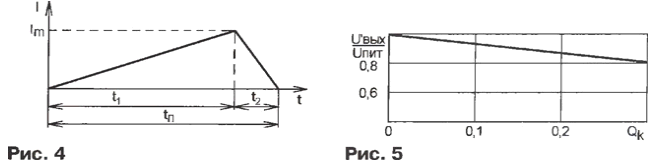
где UC2o (С22) - напряжение питания драйвера (напряжение на конденсаторе С20 или С22); UL - выходное напряжение драйвера; lL - максимальный пиковый выходной ток. В диагональ полумоста включены первичная обмотка трансформатора Т1 и дроссель L2 (индуктивность дросселя может включать в себя индуктивность рассеивания трансформатора) [4]. Трансформатор выполнен на магнитопроводе Е-Е типоразмера F-43515 фирмы Magnetics Inc. Первичная обмотка содержит 38 витков провода #19AWG, а вторичная - 5+5 витков, намотанных проводом #12AWG. Дроссель L2 наматывают на магнитопроводе F-41808EC фирмы Magnetics Inc. Обмотка дросселя L2 состоит из 8 витков провода #19AWG. Дроссель L3 выполнен на магнитопроводе тороидальной формы МРР 55930А2 фирмы Magnetics Inc. Обмотка дросселя L3 содержит 20 витков провода #12AWG. Дроссель входного фильтра L1 - Е3993 фирмы Coilcraft, его индуктивность - 900 мкГн. При включении транзистора VT1 (или VT2) через первичную обмотку трансформатора Т1 за время управляющего импульса t1 начинает протекать линейно нарастающий ток (рис. 4). Когда транзистор VT1 (или VT2) закроется, вследствие накопленной в первичной обмотке трансформатора и дросселе L2 энергии в цепи за время t2 в этом же направлении продолжает протекать линейно уменьшающийся ток. Он замыкается через диод VD7, если выключился транзистор VT1 (или через диод VD6, если выключился транзистор VT2).
Не учитывая активные потери мощности в цепи первичной обмотки трансформатора, запишем уравнения для интервалов времени t1 и t2:
где Е0 = Uпит/2 - половина напряжения питания; U'0 - выходное напряжение источника, приведенное к первичной обмотке трансформатора; L1 -суммарная индуктивность первичной обмотки трансформатора Т1 и дросселя L2. Отсюда получим выражения для времени t1 и t2 (см. рис. 4):
где lm - максимальный ток первичной обмотки трансформатора. Время протекания тока через первичную обмотку трансформатора в одном направлении tn = t1 +t2 можно выразить следующим образом:
Если принять, что
то время протекания тока равно
Из этого равенства получим уравнение для внешней характеристики источника питания. Например, для коэффициента заполнения управляющих импульсов
следует
откуда
Если обозначить
то уравнение внешней характеристики источника питания имеет вид
Внешняя характеристика блока питания показана на рис. 5. Выходное напряжение источника зависит от сопротивления резистора R17 - чем меньше сопротивление, тем меньше напряжение на выходе. Ток срабатывания защиты определяется сопротивлением датчика - резистора R5. Литература
Авторы: Р.Каров, С.Иванов, г.София, Болгария
Чувства кота, ожидаюшего возвращения хозяина
16.07.2026 Целесообразность приема пробиотиков после курса антибиотиков
16.07.2026 Резкое похудение и возврат веса могут навредить сердцу
15.07.2026
▪ ATSAMR34/35 - радио LoRa плюс микроконтроллер Cortex-M0+ для интернета вещей ▪ Сетевые A4 принтеры Xerox Phaser 3052NI и Phaser 3260DNI ▪ Лазерная катапульта для полетов на Марс ▪ Paralenz - экшн-камера для аквалангистов
▪ раздел сайта Бытовая электроника. Подборка статей ▪ статья За семью печатями. Крылатое выражение ▪ статья Как выдувают стекло? Подробный ответ ▪ статья Переключатель нагрузки от звуковых сигналов. Энциклопедия радиоэлектроники и электротехники ▪ статья Преобразователь 12/220 вольт. Энциклопедия радиоэлектроники и электротехники
Главная страница | Библиотека | Статьи | Карта сайта | Отзывы о сайте
www.diagram.com.ua |
















 Оставьте свой комментарий к этой статье:
Оставьте свой комментарий к этой статье: